This guest post comes from my fiancé who is a Psychiatrist. I've been very excited about this post for a while, because unlike me, he is a published author who has written a book on Consciousness and the philosophy of mind.
Microbes and Madness
At first glance, it would be reasonable to assume that my profession and that of the author of this fabulous blog are poles apart. However, everything in nature has a connection, and so it is not surprising to discover a fascinating area where psychiatry and bacteriology overlap.
A broad range of pathogens are known to cause psychiatric sequelae, including worms (neurocysticercosis), protozoa (cerebral malaria, toxoplasmosis), viruses (HIV, herpes simplex encephalitis, rabies), prions (Creutzfeldt-Jakob disease, kuru), and, of course, bacteria (neurosyphilis, Lyme disease, post-streptococcal syndromes). However, in the spirit of this blog, this post will be focusing on bacteria.
There are essentially four mechanisms through which bacteria cause psychiatric symptoms in humans:
I. Bacteria can infect the central nervous system and cause direct damage to brain cells.
II. Bacteria can trigger a powerful systemic inflammatory response that results in a disruption in brain function.
III. Bacteria can trigger an adaptive immune response which produces antibodies that cross-react with host central nervous system proteins.
IV. Bacteria can be the objects of a phobia.
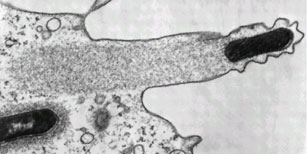
Syphilis and Lyme disease are examples of infections which involve the first mechanism. Syphilis is caused by spirochaetes of the species Treponema pallidum, and is sexually-transmitted. Lyme disease is caused by spirochaetes of the genus Borrelia, and is vector-borne, with ticks from the genus Ixodes being the commonest vector. Both diseases are associated with widespread dissemination of infection of multiple organ systems, and are notorious for their protean manifestations.
The range of possible psychiatric presentations is vast. Syphilis, in particular, can mimic any psychiatric syndrome, and was a common diagnosis in psychiatric inpatients a century ago. The possible range of presentations include delirium, dementia, psychosis, mania, and personality changes. Lesions of the frontal lobes are associated with personality changes and disinhibited behaviour, whereas those of the temporal and parietal lobes are associated with cognitive decline. Lyme disease can also mimic several different psychiatric syndromes, but typically affects the limbic system, causing disorders of emotional regulation, including panic attacks, phobias, depression, and obsessive-compulsive behaviour.
The second mechanism listed refers to sepsis-associated delirium. No human organ system is a closed system, including the central nervous system. Bacterial infections with a focus outside the outside the brain are capable of causing a systemic reaction, which affects the brain. The result is an acute confusional state, or delirium.
Common causes are pneumonias and urinary tract infections, although infections of other organ systems are also frequently implicated. Delirium presents as a transient global disorder of cognition. Typically, there is clouding of awareness, disorientation, impaired attention, fluctuating alertness with agitation or drowsiness, hallucinations, illusion, and vague delusions. The state is thought to be caused by a global disruption of brain function, which may result from the effects of a systemic inflammatory response to infection. These effects may include systemic vasodilation causing cerebral hypoperfusion, increased permeability of capillaries allowing toxins to cross the blood-brain barrier, the action of inflammatory cytokines on the brain, and increased body temperature resulting in an increase in neuronal oxygen demand.
The third mechanism is seen following infections with group A beta-haemolytic Streptococcus pyogenes, such as scarlet fever and tonsillitis. In response to infection, the adaptive immune system produces antibodies against antigens on the invading pathogen. However, some streptococcal antigens are similar in some way to antigens on host tissues, and so the antibodies produced mistakenly recognise and attack the host tissues. Examples of post-streptococcal autoimmune diseases include rheumatic fever, glomerulnephritis, and Sydenham’s chorea.
A psychiatric syndrome caused by this mechanism is PANDAS, which stands for paediatric autoimmune neuropsychiatric disorder associated with streptococcus. This typically presents as a dramatic onset of obsessive-compulsive disorder, tic disorders, or Gilles de la Tourette syndrome following an infection with group A beta-haemolytic Streptococcus pyogenes in childhood. It is thought to be a result of autoimmune damage to the basal ganglia, which is the part of the brain involved with the initiation and regulation of motor commands. Interestingly, it has also been suggested that encephalitis lethargica, a mysterious syndrome which caused an epidemic during World War I, may also be caused by a post-streptococcal autoimmune reaction.
The fourth and final mechanism listed refers to mysophobia, or the pathological fear of germs. Behavioural symptoms include repeated washing of hands, excessive cleanliness, and avoidance of social contact. Anxiety and panic attacks also occur. Although the behavioural manifestations are similar, mysophobia is not to be confused with obsessive-compulsive disorder. The former is a phobic disorder, in which the fear of germs underlies the behaviour, and the function of the behaviour is avoidance of the phobic object. In the latter, the behaviour is compulsively carried out in response to the obsession that the behaviour must be carried out.
I hope to have provided an comprehensive overview of some of the interesting ways microbes can cause mental and behavioural disturbances in humans. The function of this ability is open to speculation. The film 28 Days Later tells the story of an artificial ‘Rage’ virus. When a human is infected, he or she becomes uncontrollably aggressive, attacking other humans and infecting them with viruses in the process. Thus, the viruses’ effect on human behaviour is clearly advantageous to their spread and propagation. However, outside of fiction, the advantages of pathogens’ effects on human behaviour is less obvious. Even with rabies, on which the symptoms of the ‘Rage’ were based, there has been no documented human-to-human transmission through bites. In fact, the only documented cases of human-to-human transmission of rabies were of transplant recipients receiving corneas from infected donors! It is therefore not known what evolutionary advantage, if any, the psychiatric sequelae of infection convey to the pathogens. It is possible that they are epiphenomenal.
---
Pfister D, Siegemund M, Dell-Kuster S, Smielewski P, Rüegg S, Strebel SP, Marsch SC, Pargger H, & Steiner LA (2008). Cerebral perfusion in sepsis-associated delirium. Critical care (London, England), 12 (3) PMID: 18457586Neurosyphilis: Considerations For A Psychiatrist Mark A. Ritchie, Joseph A. Perdigao, Mark A. Ritchie
The Neuropsychiatric Assessment of Lyme Disease
A. Mazzola, G. Mazzola (2006). OCD And Beta Haemolytic Streptococcus: A Nasty Association.
Priory publishing link










































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}